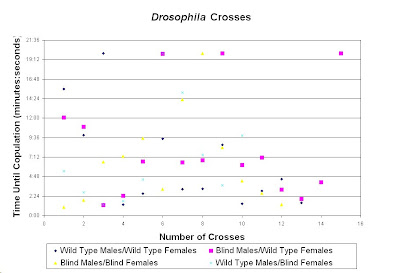
The results that are presented supports our hypothesis that pheromones, not vision, is the main element in
Drosophila melanogaster mating. Overall, the white-eyed flies and the wild-type flies mated in about the same amount of time. There was no significant difference between the four crosses. The cross with the wild type male and females had the better average time, but only a couple seconds faster than the flies with the blind males and females. There is a slight increase in the time that it took for the flies to mate when one fly was blind and the other was wild type. Therefore, no significant difference can be made from this data. Hence, it can be interpreted that the flies are able to find a mate by such means other than vision. Flies most likely find their potential mate by the detection of pheromones produced by themselves and the other sex. However, it does seem that they use more than one technique to find a mate.
Based on the results from the crosses from
Table 2 and
Table 4, it seems to be more important for the males to be able to see. The cross with the wild type males and the blind females had a faster average time than the crosses with blind males and the wild type females. This is because the males initiate the behavior and need to be able to locate the female. The females just wait until the males find them. Therefore, when the males were blind it takes a little more time for the mating to occur. Yet, the overall results show that if a fly has a mutation such as blindness, they would have no disadvantage when finding a mate. When attempting to find a mate, there was no significant difference in the fitness of the flies.
Another way of explaining the slight difference in the data is the time of day the assays were done.
Drosophila melanogaster releases their pheromones according to a circadian cycle (Stern, 2007). Since the assays were not done at the exact same time every day, there may have been more pheromones released at one particular time of day that helped guide the flies more so than any other time. There may have been contamination distributed in the collection of virgin fly vials, due to the variations in the larval stages of the flies’ maturation periods. There was also difficulty collecting adequate amounts of virgin flies, which could potentially change the results. One major variation could be the integration of non-mating assays recorded as twenty minutes, which were included in the average copulation times. All of these variations could be possible sources of error toward the experiment.
There were other similar experiments done by previous Shippensburg Students that presented conflicting data to our results. One experiment done by Nicole Thierwechter got results of blindness having a negative affect with mating (Barrett, 2008). Another experiment by Shanna Barrett had the opposite results of the males taking longer to mate in the presence of pheromones (Barrett, 2008). The results were observed in our experiment seem to be in the middle of these two experiments. In order to get a better idea of what really effects the mating of
Drosophila melanogaster more experiments would need to be done.
From the experimental data, it can be concluded that there is no significant difference between the use of vision and pheromones in
Drosophila during mating. Therefore, flies most likely find their potential mate by the detection of pheromones produced by themselves and the other sex. However, some changes could be made to better the experiment. These changes include collecting more virgin flies to prepare more mating assays, as well as conducting assays at the same time each day.
ReferencesBarrett, Shana. 2008, Nov. 30.
Effect of pheromones on mating behavior of Drosophila melanogaster.
Galino, Kathleen. 2001. A Large Family of Divergent
Drosophila Odorant-Binding Proteins Expressed in Gustatory and Olfactory Sensilla.
Genetics. 159; pp. 1059-1072.
Lin, Helping. 2005. A
Drosophila DEG/ENaC channel subunit is required for male response to female pheromones.
PNAS. 102:36; pp.831-836.
Flagg, Raymond. 2005.
Carolina Drosophila Manual 45-2620. Carolina Bioligical Supply Company.
Stern, Peter S. Yu, Lian, Choi, Man-Yeon. Jurenka, Russell A. Becker, Liron. Rafaeli, Ada. 2007. Molecular modeling of the binding of pheromone biosynthesis activating neuropeptide to its receptor.
Journal of Insect Physiology: Aug 2007; Vol. 53 Issue 8, pp. 803-818.









{kind=link}